ДРУГИ СЕМИНАРСКИ РАБОТИ
- МЕДИЦИНА: |
||||||||||||||||||||
|
||||||||||||||||||||
ГЕНСКИ МУТАЦИИ
Терминот мутации потекнува од латинскиот збор mutare-менување, а тоа се всушност наследните промени во струтурата на генскиот материјал. Промена на редоследот на нуклеотидите во ДНК кои се трајно задржуваат и се пренесуваат во наредната генерација на ќелии се викаат мутации. Мутации се одвиваат во сите организми и клетки, како нормално функционирање на организмот и во интеракција на организмот со животната средина.
Мутации претставуваат основен извор на генетската варијабилност на популацијата. Без мутации процесите на еволуција не би биле можни. Процесите на мутации се одвиваат на различно ниво на организација на наследниот материјал на организмот. Тоа е сложен процес и често има штетно него корисно дејство. Се мисли дека имаат важна улога во предизвикување на многу болести. Голем број на луѓе носат скриени мутации од рецесивен карактер, кои кога ќе се најдат во хомозиготна состојба имаат летално или штетно дејство. Учеството на мутацијата во посебни гени е различна и кај посебни човечки гени изнесува 10~5, т.е. една мутација на 100 000 делби на клетки.
Во поглед на честотата на мутирање на гените важат две правила:
1. Гените спонтано се менуваат во алелни форми со брзина константна за секој локус и таа може да е различна од еден ген до друг.
2. Мутациите често се реверзабилни А->а и а->А, а степенот на променливост во двата правци најчесто е различен.
Мутациите можат да се класифицираат според различни критериуми, но тие најчесто де делат според големината на генетскиот материјал кој е вклучен во промената и тоа на две големи групи:
■ генски мутации (или „точкасти"; „интрагенски")- различни промени во составот на нуклеотидите на поединечни гени, и
■ хромозомски мутации (или „екстрагенски"; „макромутации")- промени кои вклучуваат генски состав, сите хромозоми или пак хромозомски гарнитури.
Причинители за мутации може да бидат, на пример, грешки во текот на репликацијата на ДНК, физички застој во хромозомот или хемиска модификација голем или мал број на нуклеотиди во ДНК кои предизвикуваат различни мутагени агенси, или разни неправилности кои се случуваат во текот на процесот на создавање на гамети.
2. ГЕНСКИ МУТАЦИИ
Генски мутации подразбираат промена во самата структура на генот, тоа се најчесто промени на молекуларна основа, било да се работи за измена на распоредот на нуклеотидот или пак за инсерција на сите фрагменти во ланецот на ДНК или во внесување на нови секвенци. Промените не можат да се воочат ни во една до сега позната техника, а обично доаѓа до промена во фенотипот.
Врз основа на типот на промената која ја предизвикува еден „мутагена случка" грешките можат да се класифицираат на:
■ супституција-замена на еден нуклеотид со друг;
■ делеција-губење на еден или повеќе нуклеотида од ДНК;
■ инсерција-додавање на еден или повеќе нуклеотида во ДНК.
3. СУПСТИТУЦИОНИ МУТАЦИИСупституциони мутации можат да се поделат на транзиции и трансверзии.
А. Транзиции се супституција на еден примарен нуклеотид со друг примарен нуклеотид (C->T или T->C), или еден пурински нуклеотид со друг пурински нуклеотид (A->G или G->A). Ваквата промена, по пепликацијата на РНК, ќе доведе до замена на нуклеотидниот пар G=C со парот A=T, или обратно.
Б. Трансверзии се суптитуции на еден пурински нуклеотид со пиримидински (A->C, G->C, A->T или G->T) или обратно, пиримидински нуклеотид со пурински (C->A, T->A, C->G или T->G). Како резултат на овие промени, по репликацијата на ДНК парот A=T е заменет со парот C=G, парот T=A со парот A=T или G=C, парот G=C или A=T, а парот C=G со парот A=T или G=C. Во молекулата на ДНК можат да настанат четири вида на транзиции и осум вида на трансверзии.
4. СИНОНИМНИ И НЕСИНОНИМНИ СУПСТИТУЦИИЗамената на нуклеотидот која се одвива во кодирачките региони на гените можат да предизвикаат различни ефекти на протеините. За супституцијата се вели дека е синонимна или „тиха" (енгл. silent) бидејќи не доаѓа до промена на аминокиселините и не можат да бидат детектирани без секвенцирање на генот. Во сите останати случаи промената на нуклеотидите е несинонимна.
Несинонимни супституции, т.е. оние кои доведуваат до промена на значењето на кодонот, можат да се поделат на две групи. Тоа се смислени и бесмислени мутацииА. Смислени мутации (енгл. sense mutations) го менуваат дадениот кодон во кодон кој одредува различна аминокиселина од оние пред него што е извршена замена на нуклеотидот, на пример: GTC (валин) ->TTC (фенилаланин).
Б. Бесмислени мутации (енг. nonsense mutations) менуваат одреден кодон во еден од стоп кодоната, кое што начесто доведува до прекин во процесот на транслација, на пример: AAG (лизин) ->TAG (стоп кодон). Овие мутации лесно се откриваат бидејќи се создаваат непотполни ланци кои немаат биолошка активност.
Ако во секој чекор се оствари само една замена на нуклеотидот, тогаш секој од смислените кодони може да мутира во девет други кодони. На пример, CCU (пролин) може да мутира во три синонимни мутации (CCC, CCA и CCG) и во шест несинонимни: UCU (Ser), ACU (Thr), GCU (Ala), CUU (Leu), CAG (His), или CGU (Arg). Бидејќи универзалниот генетски код се состои од 61 х 9 = 594 можни нуклеотидни замени. Под претпоставка тие замени да се случуваат по принципот на случајност и сите кодони еднакво да бидат застапени во кодирачките региони на гените, може да се израчунаат очекуваните пропорции на различни видови на нуклеотидни супституции во генетскиот код. Поради самата структура на генетскиот код, синонимните мутации се случуваат главно на третата позиција на кодонот. Проценка на релативно учество на различни видиви на супституционални мутации во случајно генерализирани делови на гените кои ги кодираат протеините покажале дека 70% од сите можни нуклеотидни замени се на третата позиција на синонимни мутации. Од друга страна пак, сите мутации на друга позиција на кодон се несинонимни, како што е огромниот број (околу 90%) на нуклеотидни замени на прва позиција. (2)
Постојат голем број на заболувања како хемоглобинопатии чиј причинител се точкастите мутации во гените кои ги кодираат a и в синџирите на глобинот. Глобините се протеински компоненти на хемоглобинот, протеини на крвта чија функција е транспорт на киселини. Молекулот на хемоглобинот во организмот на возрасниот човек се состои од два вида a и два в синџири на глобинот и простетична група, хем. Групацијата на гените за a-глобин во геномот на човекот се наоѓаат на краткиот крак на хромозомот 16 и опфаќа два псеудогена (ipZ и ipa1) и три функционални гени: Z, кој се експримира замо во текот на првите осум недели од ембрионалниот развиток, и a1 и a2 кои се експримираат кај возрасни људи и кодираат идентични синџири на a-глобинот. Групацијата на гените за в-глобинот се наоѓа на краткиот крак на хромозомот 11 и опфаќа пет активни гени во еден псеудоген (ipP1). Од пет функционални гени кои ги кодираат глобините (Gy, Ay, е, б и в) само в-глобин експримира кај возрасен човек. До сега се опишани околу 600 точкасти мутации во гените за глобин и е расјаснета молекулската основа за поголемиот број на хемоглобинопатии.
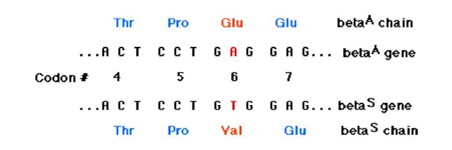
Најчесто заболување од овој вид е српеста анемија која ја предизвикува точкастата мутација во кругот на шетиот кодон на генот за в-глобин. Оваа мутација го менува кодонот GAA или GAG за глутаминската киселина (Glu) и кодон GUA или GUG за валинот (Val). Бидејќи наелектризираната аминокиселина (Glu) е заменета со неполарниот (Val), продукт на оваков мутиран ген, хемоглобин S (HbS), има изменето површинско наелектризирување и е склон кон агрегација при што еритроцитите имаат променет (српест) изглед и драстично смален век на живеење. Српеста анемија се наследува како рецесивно заболување и претставува класичен пример за мутација која во хомозиготна состојба предизвикува тешка наследна болест, додека во хетерозиготна обезбедува адаптивна предност која се состои во отпорност на паразити кои ја предизвикуваат маларијата. Една од фазите на животниот циклус на овие паразити се одвива во еритроцитите. Отпорноста на носителите на мутацијата HbS врз предизвикувачите на маларијата се темели на тоа дека овие паразити не можат да опстанат во еритроцитите кои содржат хемоглобин S.
Друга хемоглобинопатија по зачестеност е т.н. болест на хемоглобин С предизвикана од мутација на исти кодон в-глобулин како и српестата анемија, само што во овој случај кодонот GAA или GAG за глутаминска киселина (Glu) изменет во кодон AAA или AAG за лизин (Lys). (2)
Точкастите мутации кои се слуучуваат на STOP кодоните можат да го сменат STOP кодонот во кодон за некоја аминокиселина. Последица од таквите мутации е читање на генетската порака и после STOP кодонот, што е причина за синтеза за абнормално долг полипептид. Кога мутацијата ќе погоди кодон за некоја аминокиселина и го промени така да тој стане STOP кодон, последицата е прераното завршување на транслацијата, синтеза на прекраткиот и често потпуно нефункционален полипептид.
Таласемии ја сочинуваат групата на хемолитички заболувања која на молекуларна основа се одликува со неурамнотежена синтеза на a и в синџири на глобинот. Некој вид на a-таласемија настануваат поради синтеза на предолгите ланци на a-глобинот кои ги заменуваат нормалните синџири, а во клетките се присутни во многу мала количина. Синџирот на a-глобинот нормално содржи 141 аминокиселина. Но, ако STOP кодон UAA на 142 позиција мутира во кодон за глутамин (CAA), лизин (AAA), глутаминска киселина (GAA), или серин (UCA), ќе дојде до синтеза на продолжениот a-глобулински синџир, кој содржи 172 аминокиселини и е подложен на убрзана деградација, па неговата концентрација во клетката е ниска.
Бесмислени мутации исто така можат да бидат причинители за хемоглобинопатии. На пример, мутација на 145 кодон во в-глобински ген, кодон UAU или UAC за тирозин (Tyr), во STOP кодон UAA или UAG доведува до скратување на в-глобинот за две аминокиселини (нормален в-глобин содржи 146 аминокиселилни). Ваков изменет хемоглобин (т.н. McKees Rocks хемоглобин) покажува зголемен афинитет кон кис. и тешко го отпушта. За да би се компензирал овој недостаток, кај луѓето кои ја носат оваа мутација стимулирана е еритропоезата (производството на црвените крвни клетки) и зголемено лачење на еритропоетинот од бубрегот.
Како последица на бесмислени мутации настануваат и некои заболувања од групата на в-таласемија. Кај т.н. в°-талесемија, синтезата на в-глобинот потполно изостанува, па тогаш синџирите на a-глобинот се собираат во еритроидни клетки и градат преципитати кои ја оштетуваат клеточната мембрана доведувајќи ја до хемолиза и стимулација на еритропоезата. Еден облик на в°-таласемија, застапен во југоисточна Азија, настанува поради бесмислени мутации на 17 кодон во генот за в-глобинот. Оваа мутација го трансформира AAG кодонот за лизин (Liz) во STOP кодон UAG. Бидејќи мутацијата се наоѓа на самиот почеток на генот, иРНК се нестабилни, а синтезата на в-глобинот потполно изостанува.
Во практични термини забележувањето на мошне ограничена или отсутна цитоплазматична мРНК поврзано со бесмислени мутации има важна импликација за скриените мутации.
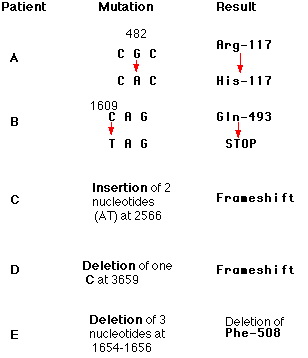
Неодамна, бесмислени мутации во фактор VIII (хемофилија А) и фибрилин (Марфанов синдром) беа поврзани со скокањето на егзон над егзон што ја продолжува муатција. Оваа беше пробано и на други гени и скокањето може да биде целосно или делумно. Механизмот на овој процес е непознат.Пример:
5. СТАПКА НА НУКЛЕИДНИ СУПСТИТУЦИИ
Стапката на нуклвотидни супституции се дефинира како број на замена на одредено место по година. Тие можат да се израчунаат со делење на бројот на нуклеотидните замени на одредено место помеѓу две хомологни реда на нуклеотиди со двојно време на дивергенција на низи кои се редат. Стапката на нуклеотидни замени можат да се процени на различни гени или помеѓу различни региони на истиот ген. Поради ова е значајно да се користат исти парови на низи иако многу често постои несигурност околу одредување на староста на многу од низите. Ако користиме исти парови на низи, тој проблем ќе се надмине.
Кај луѓето е утврдено дека скоро сите de novo мутации на гени кои предизвикуваат различни (наследни) заболувања (на пример: Apertov синдром, мултипна ендоплазија тип B и др.) настанале исклучиво во текот на гаметогенезата кај татковците. Па поради ова изгледа дека некои видови на мутации се случуваат само кај машкиот пол и исто така е поставено прашањето зошто стапката на мутации е поголема кај машките него кај жените?
Одговорот на оваа прашање лежи во фактот дека во машката герминативна линија постои значително поголем број делби на клетки него во женската. Делбата на клетките во женската герминативна линија се прекинува околу петиот месец од бременоста и мејозата се надоврзува со почетокот на пубертетот. Кај машките, делбите на клетки се континуирани и за да се појават зрели сперматозоиди треба да се случат многу делби. Ако некои видови на мутации се поврзани со делба на клетка, т.е. со репликација на ДНК, како што е случај со некои нуклеотидни замени, тогаш не се неочекувани наодите дека нивната стапка е поголема кај машките него кај жените.
До денеска се откриени голем број на наследни заболувања кај кои е утврдено дека стапката на мутации зависи од староста на таткото (нажалост, таа врска не е линеарна туку експоненцијална). Тоа се: acrodystostosis, achondroplasia, Алтеров синдром, cleidocranial dystosis, Крузонов синдром, fibrodysplasia ossificans progresiva, Марфанов синдром, окулодентодигитален синдром, Пфајверов синдром, прогерија и Варденбургов синдром.
6. ИНСЕРЦИЈА И ДЕЛЕЦИЈАМутации настанати со инсерција и делција кога ќе се додаде или одземе еден базен пар во ДНК, се менува составот и бројот на нуклеотидите, што доведува до измена во составот на иРНК, а од местото каде се вклучил погрешен кодон сите останати кодони ќе дадат погрешни аминокиселини во пептидниот синџир, така да изменетиот ген нема биолошка активност.
Инсерција и делеција најчесто се означуваат како „празнини" (енгл. gaps). Бројот на нуклеотидите кој се губи или добива може да оди од еден, или неколку, до неколку илјади.А. Исерција претставува зголемување на бројот на нуклеотидите во генот. Главен извор на исерции се т.н. мобилните генетички елементи- редови на нуклеотиди кои можат да го менуваат местото на хромозомот. Иако поретко, инсерциите можат да настанат и како последица на грешки во текот на генетската рекомбинација, а понекогаш и во текот на ДНК репликацијата. Хемиските мутагени во поголем број на случаи не придонесуват на зачестеноста на инсерциите. Исклучок се акридинските бои како агенси кои се интеркалираат помеѓу базите во ДНК. Инсерциите често доведуваат до потполно губење на функцијата на генот. Како последица на инсеција, во текот на наредната репликација на ДНК или генетички рекомбинации може да дојде до делеција на дел или цели внесени секвенци, а понекогаш и до делеција на соседни редови на нуклеотиди.
Б. Делеција претставува губење на нуклеотиди во генот. Со нив се нарушува редоследот на базите, така што транскрибираната иРНК ќе има поинакви кодони од местото на делеција или инсерција во однос на интерактните гени,а со тоа и делот на полинуклеотидната верига ќе содржи сосема различни аминокиселини.
Многу видови на веќе спомнати заболувања, a-таласемија, настануваат како последица на делеција кои ги опфаќаат a-глобинските гени. На секој хромозом 16 во геномот на човекот се наоѓаат по два гена за a-глобин, што значи дека во еритроцитите се присутни четири активни a-глобински гени. Делеција на два, три или на сите четири гена има последица во се смалено изразување на концентрација на a-глобинскиот синџир во клетките, што е пратено со поозбилни клинички симтоми на болеста.Инсерција и делециија на еден нуклеотид внатре во кодирачкиот регион на гените доведува до промена во рамките на читање на генетската порака од местото на мутација па натаму. Резултат од ваква мутација (енгл. frameshift mutation) е полипептид во кој од местото на мутацијата, па до крајот на синџирот, редоледот на аминокиселините е потполно изменет. Ваков тип на мутација е синтеза на т.н. Wayne хемоглобин, кој настанува поради делеција на третиот нуклеотид во 138 кодон (UCU) a-глобински ген. Како последица на оваа делеција се менува рамката на читање на генетски пораки и настанува a-глобин со изменет редослед на аминокиселини од 138 позиција па натаму, но и продолжен синџир, бидејќи нормалниот STOP кодон на 142 позиција (UAA) поради промени во рамките на читање се менува во кодон (AAG), а нов STOP кодон (UAG) се јавува на 147 место.
ЗАКЛУЧОКИма многу мутагенси кои вештачко го зголемуваат процентот на мутациите во организмот.
Фактори кои ги зголемуваат генетските мутации во организмот се:
- Членови на видови во одредена феографска област или етнички групи се повеќе склони кон мутации,
- Високи дози на Х-зрачење или ултра-виолетово зрачење може да ја зголеми мутацијата,
- Радиоактивни супстанци го зголемуваат процентот на мутации попретставително.
Watson и Crick покажале дека пуринските и пиримидинските бази можат да постојат во алтернативни тафтомерни форми. Аденинот и цитозинот нормално имаат NH2 група, односно се наоѓаат во амино-форма, додека гванинот и тиминот имаат кето-група СО-кето-форма. Овие форми се широко распространети во природата. Меѓутоа, од непознати причини, ретко, атомот на водород може да ја промени својата положба. со што се изразува нестабилност на овој атом, амино-формата да помине во амино-форма NH2 -> NH. Слично на тоа, кето-формата може да се промени во ретка енолна-СОН-форма. Кога ќе се јави имино или енолна форма на базите, при репликација на молекулот на ДНК ќе дојде до погрешно спарување, како и во случајот кога базите се променети под дејство на хемиски мутагени, односно на аденинот во имино форма му е комплементиран цитозинот, а на тиминот во енолна форма му одговара гванинот. Според тоа, промената на базите од една форма во друга доведува, исто така, до мутации од типот на супстанца, што е означено како спонтани мутации бидејќи не е познато кој фактор доведува до овие промени. Инаку оваа појава ретко се случува во природата.
KOPИCTEHA ЛИТЕРЛТУРА
1. Kičić M, Krajinčanić B. Medicinska genetika. Defektološki Fakultet Univerziteta u Beogradu, Beograd, 1994.
2. Tucić N, Matić G. O genima i ljudima. Centar za primenjenu psihologiju, Beograd,
2002.
3. Митева H. Општа биологија. Вест, Скопје, 1998.
4. Remoin DL, Connor JM, Pyeritz RE. Emery and Rimoin's principles and practice of Medical Genetics. Vol I, Third edition, Churcill Livingstone, New York, 1997.
5. Mutatins (http://www.biology-online.org/2/8_mutatins.html)
6. Mtatinon Frequency and Poliploidy (http://www.biology_online.org/2/9_mutations.html
Download
СЕМИНАРСКА РАБОТА у wordu » » »

Besplatni Seminarski Radovi - Бесплатно семинарска работа