
OSTALI SEMINARSKI RADOVI
IZ HEMIJE: |
|||||||||||||||||||||||||||||||
|
|||||||||||||||||||||||||||||||
ENZIMI
1. Preciscavanje enzima
Enzimi se u industriji slično kao u laboratoriji prečišćavaju hromatografskim metodama. Za prečišćavanje enzima koriste se uglavnom tri tipa hromatografskog prečišćavanja:
- jonoizmenjivačka hromatografija
- gel-permeabilna hromatografija
- afinitetna hromatografija
2. Priprema enzima za prodaju
Nakon prečišćavanja enzima i koncentrovanja često se dobija viša aktivnost enzima od one koja je potrebna u industriji. Osnovni problem je održati aktivnost enzima duže vreme. Da bi se to postiglo neophodno je stabilizovati enzim. Industrijski enzimi sadrže relativno malu količinu aktivnog enzima (manje od 10 %). Ostatak čine neaktivni proteini, stabilizatori, soli i drugo. U cilju stabilizacije enzima primenjuju se tri pristupa:
- dodavanje aditiva
- kovalentna modifikacija proteina i
- imobilizacija enzima
Kao aditivi mogu se primeniti neutralne soli kao što su amonijum-sulfat i kalijum hidrogenfosfat koji povećavaju interakcije između hidrofobnih delova enzima i na taj način ga stabilizuju. Visoka koncentracija NaCl sprečava rast mikro-organizama. Pored toga koriste se nisko molekularni polioli (glicerol, sorbitol i manitol) koji sprečavaju rast mikroorganizama. Glicerol takođe ima ulogu za stabilizaciju enzima na niskim temperaturama jer sprečava formiranje kristala leda. Pored toga ukoliko se isporučuje vodeni rastvor enzima dodaju se antibiotici i estri benzoeve kiseline kao prezervativi.
Enzimi su znatno stabilniji u suvom stanju nego u rastvoru. Stoga se mnogo češće enzimi isporučuju u suvom obliku. Za prevođenje enzima u suvi oblik primenjuju se dva postupka:
- liofilizacija i
- sprej sušenje
Drugi postupak se zbog ekonomičnosti u industriji mnogo češće koristi.
U rastvor enzima se dodaju skrob, laktoza, karboksi metilceluloza i polielektroliti
koji povećavaju stabilnost enzima prilikom sušenja.
Sprej sušenje je postupak koji podrazumeva atomizaciju rastvora pomoću
rasprskivača. Mešanjem sa toplim vazduhom dolazi do isparavanja tečnosti
i izdvajanja čvrstog proizvoda.
3. Tehnike imobilizacije enzima
Imobilizovani enzimi su nerastvorni enzimi. U reakcijama koje se izvode
u vodenoj sredini enzimi se mogu učiniti nerastvornim umrežavanjem ili
fiksiranjem enzima za čvrsti nosač. U organskoj sredini mogu se koristiti
suspenzije liofilizovanih enzima ili enzimi fiksirani za nosač.
Metode imobilizacije
Enzim se može imobilizovati na nekoliko načina:
- adsorpcijom
- kovalentnim vezivanjem
- zarobljavanjem unutar umreženog polimera
- zarobljavanjem unutar semipermeabilne membrane
Adsorpcija enzima na pogodnom nosaču je jednostavna metoda pogodna
za nanošenje velike količine enzima (1 g po gramu matriksa). Izvodi se
mešanjem enzima sa adsorbentom pri odgovarajućim uslovima (pH, jonska
sila), inkubiranjem i na kraju ispiranjem enzima koji se nije vezao. Adsorpcija
je kombinacija hidrofobnih interakcija i jonskih interakcija koje potiću
od građenja soli između AK i nosača. Bitno je da u toku reakcije ne dolazi
do slabljenja ovih adsorpcionih veza usled promene pH, vezivanja supstrata
ili ‘promene jonske sile. Pogodni adsorbenti koji se najčešće koriste
su: jonoizmenjivači (DEAE Sephadex i CM Sephadex), porozni ugljenik, gline,
staklo, hidratisani metalni oksidi i polimerni aromatični ugljovodonici.
Iako su jonoizmenjivači najskuplji njihova primena je opravdana jer se
mogu koristiti više puta pošto nakon dezaktivacije enzima mogu da se regenerišu
spiranjem enzima koncentrovanim rastvorom soli.
Imobilizacija enzima kovalentnim kuplovanjem za nerastvorni matriks
je intenzivno primenjivana u istraživačkim laboratorijama. Nedostatak
ove metode je relativno mala količina enzima koji se vezuje (0,02 grama
po gramu matriksa). Prednost ove metode je jako vezivanje enzima i malo
curenje enzima sa nosača. Najpogodnija grupa u enzimu koja se vezuje je
amino ostatak lizina, mada se veza može ostvariti i preko sumpora iz cisteina
ili kiseonika iz serina i treonina. Prednost lizina je što se on retko
nalazi u aktivnom centru.
Zarobljavanje enzima unutar umreženog polimera moguće je izvesti
na dva načina. Prvi način podrazumeva aktiviranje polimera (npr. acilovanjem
amina sa hloridom akrilne kiseline) a zatim kopolimerizacijom sa akrilamidom
i bis akrilamidom pri čemu nastaje gel. Drugi način podrazumeva zarobljavanje
u umreženim polimerima (rastvor natrijum-alginata i enzima se sipa u rastvor
kalcijum-hlorida). Na ovaj način se stvaraju zrna kalcijum-alginata u
kojima je zarobljen enzim. Ova metoda je podesnija za zarobljavanje delova
ćelija ili samih ćelija nego enzima. Nedostatak je što veći supstrati
ne mogu da priđu enzimu.
Imobilizacija u semipermeabilnim membranama se može izvesti na
dva načina. Jedan je ugrađivanje polimera u membranu (u rastvor sebacoil-hlorida
u organskom rastvaraču dodaje se vodeni rastvor heksametilen-diamina i
enzima) pri čemu se oko enzima stvara semipermeabilna membrana koja dozvoljava
supstratu i proizvodu da prolaze kroz nju dok enzim ne može da izađe.
Drugi sistem su tzv. hollow fibre ili šuplja vlakna od semiperme-abilnih
membrana kroz koja se u jednom pravcu propušta enzim a u drugom supstrat.
Enzim ne može da prođe kroz membranu i da pređe u deo u kome se nalazi
supstrat.
4. Enzimi u industriji detergenata
Industrija detergenata troši negde oko 25 do 30 % proizvedenih enzima.
Nečistoće koje se javljaju mogu biti po svom hemijskom sastavu ugljeni
hidrati, proteini i lipidi. Primena enzima za razlaganje ovih jedinjenja
omogućava kraće vreme i nižu temperaturu prilikom pranja. Od enzima koriste
se amilaze, proteaze i u nekim novijim formulacijama detergenata lipaze.
Udeo sirovog enzima kreće se između 0,4 i 0,8 %. Enzimi se isporućuju
kao granulat (prečnika 0,5 mm) pri čemu su oni sami zaštićeni slojem soli
(NaCl) i šećera. Pored toga dodaje se i karboksimetilceluloza koja gradi
koloid.
Proteaze (alkalaza (B. licheniformis) i maksataza (B. licheniformis)
na nizem pH i savinaza (B. amyloliquefaciens) i esperaza (B. licheniformis)
na visem pH); Amilaze (Termamil (B. licheniformis)).
5. Enzimi u industriji hrane
Proteaze u industriji hrane se uglavnom primenjuju u proizvodnji sireva
renin ili himozin i u postupku osvežavanja mesa papain.Renin
se izoluje iz predželuca teleta mada je danas ovaj postupak zamenjen proizvodnjom
rekombinantnih mikroorganizama koji ga luče. Renin je selektivna proteaza
koja hidrolizuje vezu između fenilalanina i metionina.
Hidrolizom kazeina narušava se njegov stabilizujući koloidni efekat što
rezultuje zgrušavanjem mleka.
Meso starijih životinja je obično žilavo. Ono se može omekšati ukoliko
se životinji pre klanja u vratnu venu ubrizga neaktivni papain. Papain
se izoluje iz lateksa papaje. Dezaktiviranje se postiže redukcijom enzima
odnosno građenjem disulfida. Prilikom klanja životinje enzim se aktivira
zbog oksidacionih uslova. Ovaj enzim hidrolizuje kolagen i na taj način
omekšava meso čineći ga pogodnim za prodaju.
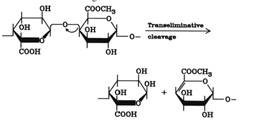 Pektini
koji se nazivaju i galakuronani ili ramnogalakturonani su heteropolisaharidi
koji se sastoje iz delimično esterifikovanih metanolom galakturonskih
jedinica. Lanci poligalakturonske kiseline su povezani ramnozom. Pektin
je sastavna komponenta ćelijskog zida. Prisustvo pektina predstavlja ozbiljan
problem u postupku bistrenja sokova i vina i otežava ceđenje. Pektinaze
iz Aspergilus nigera razgrađuju ćelijski zid i omogućavaju taloženje pektina
i njihovo uklanjanje. Pektinaze predstavljaju smešu više enzima sa različitim
mehanizmom delovanja (pektin metilesteraza hidrolizuje estarsku vezu,
pektindeacetilesteraza hidrolizuje acetilostatak i pektin depolimeraza
koja hidrolizuje vezu između dve galakturonske jedinice.
Pektini
koji se nazivaju i galakuronani ili ramnogalakturonani su heteropolisaharidi
koji se sastoje iz delimično esterifikovanih metanolom galakturonskih
jedinica. Lanci poligalakturonske kiseline su povezani ramnozom. Pektin
je sastavna komponenta ćelijskog zida. Prisustvo pektina predstavlja ozbiljan
problem u postupku bistrenja sokova i vina i otežava ceđenje. Pektinaze
iz Aspergilus nigera razgrađuju ćelijski zid i omogućavaju taloženje pektina
i njihovo uklanjanje. Pektinaze predstavljaju smešu više enzima sa različitim
mehanizmom delovanja (pektin metilesteraza hidrolizuje estarsku vezu,
pektindeacetilesteraza hidrolizuje acetilostatak i pektin depolimeraza
koja hidrolizuje vezu između dve galakturonske jedinice.
6. Enzimi u proizvodnji visoko-fruktoznog sirupa
Skrob se može hidrolizovati do glukoze kiselom
hidrolizom. Ovaj postupak je danas u večini zemalja zamenjen enzimskim
postupkom. U prvoj fazi se skrob prevodi u rastvor. 30 % suspenzija skroba
se uz mešanje zagreva do 100 do 105 ° C. Nakon toga se dodaje termostabilna
bakterijska a-amilaza koja hidrolizuje a-1,4-glikozidne veze (ne hidrolizuje
a-1,6-glikozidne veze). Nakon toga se rastvor delimično hidrolizovanog
skroba ohladi i smanji se pH vrednost na 4 do 4,5 pri čemu se deaktivira
amilaza. Zatim se dodaje glukoamilaza izolovana iz Aspergillus nigera.
Glukoamilaza hidrolizuje oligosaharide sa neredukujučeg kraja i ponovo
hidrolizuje samo 1,4 glikozidne veze. Na ovaj način se dobija rastvor
koji sadrži 97 % glukoze i 3 % oligosaharida. Po završetku reakcije enzim
se odvaja na jonoizmenjivaču a glukoza se izdvaja uparavanjem.
U Americi se veliki deo ovako proizvedenog glukoznog sirupa prevodi u
visoko fruktozni sirup. Ovaj sirup se koristi kao zaslađivač jer je ekvivalentan
po slatkosti saharozi. Pre toga je iz sirupa neophodno udaljiti kalcijum
koji se dodaje za poboljšanje rada amilaze. Nakon toga dodaje se glukoizomeraza.
Ovaj enzim konvertuje ovakav rastvor u smesu koja sadrži 42 do 46 % fruktoze
(ostalo je glukoza). U ovu svrhu se danas koriste imobilizovani enzimi
(u kolonama). 1 kg imobilizovanog enzima proizvodi 10 do 11 tona sirupa.
Čista fruktoza se može izolovati hromatografijom na zeolitima ili kalcijumovim
solima katjonskih jonoizmenjivačkih smola.
U Evropi kao alternativa fruktoznom sirupu ranije opisanom koristi se
invertni šećer čija je najvažnija prednost u odnosu na saharozu manja
tendencija kristalizacije. Za hidrolizu saharoze do glukoze i fruktoze
može se koristiti enzim invertaza izolovan iz kvasca.
7. Hromatografske metode u preciscavanju enzima
Enzimi se u industriji slično kao u laboratoriji prečišćavaju hromatografskim metodama. Za prečišćavanje enzima koriste se uglavnom tri tipa hromatografskog prečišćavanja:
- jonoizmenjivačka hromatografija
- gel-permeabilna hromatografija
- afinitetna hromatografija
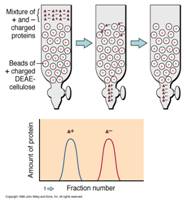 Jonoizmenjivačka
hromatografija zasniva se na činjenici da su proteini u zavisnoti
od pH vrednosti različito naelektrisane čestice odnosno iznad izoelektrične
tačke su negativno naelektrisani a ispod izoelektrične tačke su pozitivno
naelektrisani. Ovakva njihova osobina im omogućava da se vezuju za jonoizmenjivače.
U industriji su kao jonoizmenjivaći korišćeni DEAE celuloza (dietilaminoetil-celuloza)
kao anjonski jonoizmenjivač i CM celuloza (karboksimetil-celuloza) kao
katjonski jonoizmenjivač. Danas su ovi izmenjivači uglavnom zamenjeni
sa SP i QAE odnosno sulfopropil i kvaternernim amino etil derivatima.
U industriji se često primenjuje takozvani "bač" postupak u
kome se jonoizmenjivač meša sa rastvorom enzima a zatim se supernatant
u kome se nalaze proteini koji nisu vezani za kolonu odvaja centrifugiranjem.
Izmenjivač sa vezanim proteinima se zatim prebacuje u kolonu i proteini
se eluiraju promenom jonske sile rastvora ili promenom pH rastvora.
Jonoizmenjivačka
hromatografija zasniva se na činjenici da su proteini u zavisnoti
od pH vrednosti različito naelektrisane čestice odnosno iznad izoelektrične
tačke su negativno naelektrisani a ispod izoelektrične tačke su pozitivno
naelektrisani. Ovakva njihova osobina im omogućava da se vezuju za jonoizmenjivače.
U industriji su kao jonoizmenjivaći korišćeni DEAE celuloza (dietilaminoetil-celuloza)
kao anjonski jonoizmenjivač i CM celuloza (karboksimetil-celuloza) kao
katjonski jonoizmenjivač. Danas su ovi izmenjivači uglavnom zamenjeni
sa SP i QAE odnosno sulfopropil i kvaternernim amino etil derivatima.
U industriji se često primenjuje takozvani "bač" postupak u
kome se jonoizmenjivač meša sa rastvorom enzima a zatim se supernatant
u kome se nalaze proteini koji nisu vezani za kolonu odvaja centrifugiranjem.
Izmenjivač sa vezanim proteinima se zatim prebacuje u kolonu i proteini
se eluiraju promenom jonske sile rastvora ili promenom pH rastvora.
Gel permeabilna hromatografija se zasniva na odvajanju proteina na osnovu njihove molekulske mase. Koriste se matriksi dobiveni umrežavanjem dekstrana (Sephadex G), poliakrilamida (Bio Gel P), agaroze (Sepharose C) i drugi. Svi ovi matriksi izrađuju se sa različitom poroznošću i nose oznake 25, 50 , 100 i 200, na primer, što označava veličinu pora. Što je veća veličina pora manja je rigidnost polimera i sa njim je teže raditi. Razvijeni su noviji znatno skuplji polimeri koji daju bolje rezultate (smesa poliakrilamida i agaroze i kopolimer etilenglikola i metakrilata). Nedostatak metode je razblaživanje uzorka.
Afinitetna hromatografija se zasniva na vezivanju proteina za imobilizovani
specifični ligand za taj protein. Ovaj tip hromatografije može dati najbolje
rezultate u prečišćavanju proteina. Kao nosač najčešće se koristi agaroza
za koju se ligand ili antitelo vezuje kovalentnom modifikacijom. Ovakvi
matriksi su skupi i primenjuju se samo ukoliko postoji ekonomska opravdanost
(skup enzim, veliki broj drugih proteina sličnih osobina). Eluiranje se
izvodi rastvorom liganda koji je vezan za matriks.
8. Faktori koji uticu na primenu enzima
Uticaj pH
Enzimi su kao i drugi proteini amfoterni molekuli odnosno na njihovoj
površini se nalaze kisele i bazne aminokiseline koje u zavisnosti od pH
vrednosti mogu biti naelektrisane.Kada je ukupno naelektrisanje enzima
0 enzim se nalazi na izoelektričnoj tački i tada ima najmanju rastvorljivost.
Pored toga naelektrisane AK su odgovorne za vezivanje kofaktora i njihova
neutralizacija utiće na vezivanje metalnih jona. Pored toga naelektrisane
AK u aktivnom mestu mogu biti odgovorne za izvođenje hemijske reakcije.
Stoga svaki enzim ima optimalnu pH vrednost na kojoj se odvijaju reakcije.
Na izuzetno niskoj i visokoj pH vrednosti dolazi do denaturacije proteina
odnosno narušavanja strukture proteina.
Uticaj temperature
Slično kao i kod drugih hemijskih reakcija i enzimske reakcije se ubrzavaju
sa porastom temperature. Međutim za razliku od klasičnih reakcija sa porastom
temperature dolazi do denaturacije proteina. Stoga se za svaki enzim proverava
termostabilnost odnosno određuje se optimalna temperatura za izvođenje
hemijske reakcije. Prilikom određivanja mora se voditi računa i o vremenu
trajanja reakcije jer prilikom produženog izlaganja enzima dolazi do denaturacije
iako je enzim stabilan na toj temperaturi.
Inhibicija enzima
Mnoge supstance mogu delimično ili potpuno smanjiti aktivnost enzima.
Neke od njih deluju nespecifično i to je slučaj sa reagensima koji izazivaju
denaturaciju (urea,
Druge supstance koje se na specifičan način vezuju za enzim nazivaju se
inhibitori. Inhibicija može biti reverzibilna (aktivnost enzima se vraća
po uklanjanju inhibitora iz sistema) ili ireverzibilna (joni olova i žive
se vezuju jakim vezama za aminokiselinski kostur i zazivaju ireverzibilnu
inhibiciju).
9. Enzimi u sintezi aminokiselina
Postoji nekoliko različitih pristupa za enzimsku sintezu aminokiselina.
Prvi pristup za sintezu L-aminokiselina zasniva se na selektivnoj hidrolizi
N-acilaminokiselina. Racemska smesa aminokiselina se aciluje i zatim hidrolizuje
na pH 8,5 dejstvom aminoacilaze iz Aspergilus 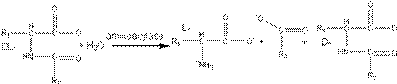 oryzae.
Hidrolizuje samo L-derivat dok D-derivat ostaje nehidrolizovan. Racemizacijom
D-derivata i ponovnom hidrolizom može se povećati prinos L-aminokiseline.
Koristi se imobilizovani enzim na DEAE-Sephadexu.
oryzae.
Hidrolizuje samo L-derivat dok D-derivat ostaje nehidrolizovan. Racemizacijom
D-derivata i ponovnom hidrolizom može se povećati prinos L-aminokiseline.
Koristi se imobilizovani enzim na DEAE-Sephadexu.
Reakcijom aldehida sa HCN i amonijum-karbonatom nastaje racemski hidantoin.
Hidantoin se primenom enzima hidantoinaze može selektivno prevesti u L-
ili D-amino-kiselinu. Hidantoinaza se izoluje iz Arthrobacter-a
(L-derivati) ili Pseudomonas striata (D-derivati). D-aminokiseline
su značajne u sintezi antibiotika i insekticida.
Asparaginska kiselina je važan intermedijer u sintezi aspartama (veštački
zaslađivač). Ova kiselina može se sintetisati enzimski iz fumarne kiseline.
U sintezi aspartama primenjuje se enzim termolizin koji daje samo a-aspartam
odnosno željeni proizvod.
preuzmi
seminarski rad u wordu » » »