OSTALI SEMINARSKI RADOVI
IZ BIOLOGIJE : |
|||||||||||||||||||||||||||||||||||||||||||
|
|||||||||||||||||||||||||||||||||||||||||||
HROMOZOMI
Hromozomi su tjelašca (grč. hromos – boja, soma – tijelo) karakterističnog oblika koja se u jedru eukariotskih organizama mogu uočiti za vrijeme diobe. Zahvaljujući tome što se dobro boje, hromozomi se mogu u određenoj fazi diobe posmatrati pod svjetlosnim mikroskopom. Najbolje se uočavaju za vrijeme metafaze mitoze pa se nazivaju metafazni hromozomi. Najveći dio DNA kod eukariota smješten je u hromozomima, samim tim i najveći dio gena, pa su hromozomi najvažniji nosioci nasljednosti u ćeliji.
Građa metafaznog hromozoma
Svaki metafazni hromozom se sastoji od:
- Dvije sestrinske hromatide koje sadrže po jedan molekul DNA i s obzirom
da nastaju replikacijom, ti molekuli su potpuno jednaki po sadržaju gena,
zato se hromatide nazivaju sestrinske.
- Centromere ili primarnog suženja koje spaja hromatide.
Hromatide
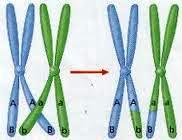 Hromatida
je uzdužna polovina metafaznog hromozoma. Prije nego
što se izvrši replikacija DNA, hromozom ima jedan molekul DNA koji se
povezuje sa proteinima i nagradi jednu hromatidu. Poslije replikacije
hromozom dobija još jedan potpuno identičan molekul DNA (hromatidu), zahvaljujući
semikonzervativnosti replikacije.
Hromatida
je uzdužna polovina metafaznog hromozoma. Prije nego
što se izvrši replikacija DNA, hromozom ima jedan molekul DNA koji se
povezuje sa proteinima i nagradi jednu hromatidu. Poslije replikacije
hromozom dobija još jedan potpuno identičan molekul DNA (hromatidu), zahvaljujući
semikonzervativnosti replikacije.
Hromatide mogu biti:
- Sestrinske, to su hromatide jednog hromosma, njihove uzdužne
polovine spojene centromerom; prije nego se izvrši krosing – over ove
hromatide su potpuno međusobno jednake jer nastaju replikacijom.
- Nesestrinske, to su međusobno hromatide para homolognih hromozoma
i u pahitenu profaze mejoze i između njih se razmjenjuje genetički materijal
(krosing – over).
Krosing-over je razmena genetičkog materijala između nesestrinskih hromatida homologih hromozoma. U pahitenu profaze mejoze i dolazi do obrazovanja hijazmi (mostića) između nesestrinskih hromatida, a zatim do prekida na tim mestima i međusobne razmjene odgovarajućih delova – krosing-overa.
Broj hromatida u somatskim ćelijama
Normalne tjelesne ćelije diploidnih organizama uvijek sadrže parove homolognih
hromozoma. Sve ove ćelije su u principu iste kao zigot (oplođena jajna
ćelija) od koga su nastale mitotičkim diobama. U paru homolognih hromozoma
jedan hromozom potiče iz očeva (spermatozoida), a drugi iz majčine garniture
(jajne ćelije). Zbog krosing – overa koji se dešava u mejozi I pri obrazovanju
polnih ćelija, hromozomi koji su naslijeđeni nisu jednaki majčinim ili
očevim. Broj hromatida u somatskoj ćeliji zavisi od faze ćelijskog ciklusa
u kojoj se ćelija trenutno nalazi. U najkraćem ćelija prije diobe duplira
količinu DNA, broj hromatida, da bi ih ravnomjerno raspodijelila na kćeri
ćelije za vrijeme diobe.
Broj hromatida u gametima
Polne ćelije, gameti, sadrže samo po jedan hromozom iz svakog para. U
mejozi II, poslije koje nastaju gameti, dolazi do podjele hromozoma na
hromatide koje tada postaju novi hromozomi. Tako, hromatide polnih ćelija
imaju jedan molekul DNA.
Broj hromatida u toku ćelijskog ciklusa
Broj hromatida je promjenjiv u toku ćelijskog ciklusa. Pošto se replikacija
dešava za vrijeme S faze interfaze ćelijskog ciklusa; hromozom se sastoji
od :
- Jedne hromatide u periodu od anafaze mitoze pa do S faze narednog ćelijskog
ciklusa ;
- Dvije hromatide od S faze do anafaze mitoze.
Posmatrano na ukupan broj hromozoma humana tjelesna ćelija sadrži:
- 46 hromatida, odnosno, onoliko hromatida koliko i hromozoma, u periodu
od anafaze mitoze pa do S faze narednog ćelijskog ciklusa;
- 92 hromatide, u period od S faze interfaze pa do anafaze mitoze.
Drugačije rečeno, u ćeliji se prije diobe broj hromatida duplira da bi
se mitozom ravnomjerno podijelile novonastalim kćerkama ćelijama. Veoma
je važno da novonastale ćelije dobiju isti genetički materijal kao što
je imala ćelija od koje su nastale.
Centromera
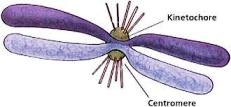 Centromera
je primarno suženje na hromozomu koje spaja sestrinske hromatide.
Na centromeri se nalazi proteinska struktura, kinetohor, koja se formira
na početku mitoze i ima ulogu da veže hromozom za diobeno vreteno. Ona
usmjerava kretanje hromozoma za vrijeme diobe pošto se za nju vezuju konci
diobenog vretena.
Centromera
je primarno suženje na hromozomu koje spaja sestrinske hromatide.
Na centromeri se nalazi proteinska struktura, kinetohor, koja se formira
na početku mitoze i ima ulogu da veže hromozom za diobeno vreteno. Ona
usmjerava kretanje hromozoma za vrijeme diobe pošto se za nju vezuju konci
diobenog vretena.
U anafazi mitoze (mejoze II) se uzdužno dijeli, čime se sestrinske hromatide
razdvajaju ( sada su to novi hromozomi) i odlaze na suprotne polove ćelije.
Zahvaljujući tome novonastale, kćerke ćelije dobijaju međusobno iste gene
i istovremeno iste gene kao majka ćelija od koje su diobom nastale. Nepravilna,
poprečna podjela centromera dovodi do duplikacija i delecija (to su vrste
strukturnih aberacija hromozoma).
Centromera dijeli hromozom na dva kraka:
- Kratki krak (prema međunarodnom dogovoru obilježava se sa r)
je dio hromosma iznad centromere;
- Dugi krak (obilježava se sa q) je dio hromozoma ispod centromere.
Pored ovih dijelova, specifičnih za sve hromozome, poneki hromozomi mogu
imati i sekundarno suženje. Akrocentični hromozomi u humanom kariotipu
imaju to suženje.
Kinetohor
Kinetohor je parna proteinska struktura u predjelu centromere hromozoma.
Svaka hromatida metafaznog hromozoma ima jedan kinetohor. Za njega se
vezuju niti diobenog vretena pa je svaki hromozom u metafazi mitoze povezan
za oba pola ćelije. Hemijski sastav kinetohora nije sasvim tačno utvrđen
mada se sigurno zna da sadrži bazne proteine. Uočen je elektronskim mikroskopom
i različito je građen u hromozomima različitih organizama:
- Kod viših biljaka je loptastog oblika
- Kod čovjeka, različitih životinja i nižih biljaka je troslojan.
Troslojni kinetohor se sastoji od 3 dijela:
- Dio za koji su pričvršćene niti diobenog vretena i on je najgušći
- Srednji dio koji je blijeđi
- Unutrašnji, gušći dio koji naliježe na hromatin.
Vrste hromozoma
Prema položaju centromere u hromozomu razlikuju se:
- Metacentrični hromozomi, kod kojih je centromera postavljena
medijalno, a p i q kraci su približno iste dužine. U ovu grupu spadaju
hromozomi 1, 3, 16, 19 i 20. par hromozoma u humanom kariotipu.
- Submetacentrički hromozomi, kod kojih centromera nije
smještena na sredini, nego je pomjerena ka jednom kraju i dijeli hromozom
na dva kraka nejednake dužine (kraći p krak i duži q krak). Submetacentrički
hromozomi u kariotipu čovjeka su 2, 4, 5, 6, 7, 8, 9, 10, 11, 12, 17,
18 i X hromozom. Žene imaju paran, a muškarci zbog prisustva jednog X
hromosma, neparan broj submetacentričkih hromozoma u kariotipu.
- Akrocentrički hromozomi su oni hromozomi kod kojih
se centromera nalazi sasvim blizu jednog kraja hromozoma tako da dijeli
hromozom na sasvim kratak p krak i mnogo duži q krak. Akrocentrički hromozomi
u kariotipu čovjeka su: 13, 14, 15, 21, 22 i Y hromozom. Muškarci, zbog
prisustva Y hromozoma, imaju neparan broj akrocentričkih hromozoma u tjelesnim
ćelijama, a žena ima paran broj. Spermatozoid muškarca može da sadrži
različit broj akrocentričkih hromozoma: 5 (ako nema Y hromozom) ili 6
(ako sadrži Y hromozom).
- Telocentrički hromozomi, kod kojih je centromera postavljena
na samom kraju hromozoma, odlikuje ih odsustvo p kraka, nema ih u humanom
kariotipu.
Homologni hromozomi
Homologni hromozomi su spareni hromozomi identični po veličini, obliku i funkciji. U svakom paru homolognih hromozoma (bivalent ili tetrada) jedan potiče iz majčine, a drugi iz očeve garniture hromozoma. Sparivanje homolognih hromozoma dešava se u zigotenu profaze mejoze I i naziva sinapsa, a razdvajanje se vrši u anafazi mejoze I. Između sparenih hromozoma dolazi do krosing – overa u pahitenu profaze mejoze I.
Broj hromozoma
Broj hromozoma je stalan i karakterističan za svaku biološku vrstu i
naziva se kariotip. Tjelesne (somatske) ćelije imaju diploidan (grč. diploos
– dvostruk) broj hromozoma, obilježava se kao 2n. Tjelesna ćelija čovjeka
ima 46 hromozoma ili dvije garniture po 23 hromozoma, pri čemu jedna garnitura
potiče od majke, a druga od oca pa se tako obrazuju 23 para homolognih
hromozoma.
Polne ćelije ili gameti (kod čovjeka su to spermatozoidi i jajna ćelija)
sadrže upola manji broj hromozoma u odnosu na tjelesne ćelije, nazvan
haploidan (grč. haploos – jednostruk), obilježava se kao n. Ako tjelesna
ćelija ima dvije, onda će polna ćelija imati jednu garnituru hromozoma.
Broj hromozoma u polnim ćelijama čovjeka je 23.
Hromatin
Hromatin se uočava u interfaznom jedru (to je jedro ćelije koje se nalazi između dva dijeljenja). On je u obliku hromozomskih vlakana. Tokom ćelijske diobe dolazi do kondenzovanja (spiralizacije) hromatinskih vlakana tako da ona postaju samostalna tjelašca – hromozomi. Hromatin se sastoji od DNA, male količine RNA i dvije vrste proteina: histona i nehistonskih proteina. Histoni su bazni proteini zbog prisustva veće količine baznih aminokiselina (najviše lizina i arginina). Razlikuje se pet klasa (vrsta) histona koji se obilježavaju kao H1, H2A, H2B, H3 i H4. Ovi proteini su veoma slični kod različitih eukariotskih organizama, što znači da se tokom evolucije živog svijeta nisu mnogo mijenjali pa se za takve proteine kaže da su konzervativni (očuvani). Oni su gradivni proteini hromatina jer učestvuju u pakovanju DNA, koja se oko njih namotava (kao konac oko kalema) da bi se ogromna dužina DNA mogla smjestiti u sićušno jedro. Zato se hromatin vidi kao perlasta struktura, gdje su perle, ustvari, histoni oko kojih je namotana molekula DNA. Nehistonski proteini su kiseli proteini hromatina i broj vrsta nehistonskih proteina je mnogo veći od histonskih (40-80 različitih vrsta). U njih spadaju mnogobrojni enzimi koji u jedru učestvuju u procesima replikacije i transkripcije. Ako su histoni gradivni onda su nehistonski proteini funkcionalni proteini hromatina. Prema jačini bojenja razlikuju se dvije vrste hromatina: heterohromatin i euhromatin. Heterohromatin je kondenzovan (spiralni navoji DNA koji su zbijeni) pa je zbog toga tamnije obojen i lako se uočava u jedru. Istovremeno je to vrsta hromatina koja je neaktivna u transkripciji. Euhromatin je difuzan (rasplinut – manje su zbijeni navoji DNA) pa je uslijed toga svjetlije obojen. Ćelije koje intenzivno sintetišu neki protein imaju malo heterohromatina, a dosta euhromatina i njihova jedra su svjetlija.
Hromozomske aberacije
Hromozomske aberacije (lat. aberatio = odstupanje
od normalnog) su promjene u genomu koje obuhvataju cijele
hromozome ili njihove dijelove. Mogu se uočiti pod mikroskopom za razliku
od genskih mutacija. U zavisnosti od toga da li predstavljaju promjene
u građi ili broju hromozoma razlikuju se dva tipa:
- Strukturne aberacije (promjene u građi hromozoma)
- Numeričke aberacije (promjene u broju hromozoma)
Strukturne aberacije hromozoma
Ovaj tip aberacija je posljedica prekida i poremećaja linearnog kontinuiteta
hromozoma. Prekidi se nekada javljaju spontano, ali mogu biti i izazvani
različitim mutagenima. Hromozomi se mogu prekidati na bilo kom stadiju
ćelijskog ciklusa. Prekinuti dijelovi, pošto su izgubili telomere, teže
da se međusobno spoje, a da bi se to desilo hromozomi moraju da budublizu
jedan drugoga. Na osnovu toga kako se prekinuti dijelovi povezuju (preraspoređuju)
razlikuju se četiri osnovna tipa strukturnih aberacija:
- Delecije
- Duplikacije
- Inverzije
- Translokacije.
Duplikacije i delecije
Duplikacije predstavljaju udvajanje, a delecije gubljenje dijelova hromozoma. Uzrok ovih aberacija je vrlo često nejednak (nepravilan) krosing-over. Do njega dolazi kada se nesestrinske hromatide homologih hromozoma ne postave simetrično naspramno jedna drugoj. Rezultat takvog krosing-overa su hromatide kod kojih se na jednoj javlja dupliran (udvostručen) deo, a na drugoj hromatidi taj deo nedostaje (delecija) – vidi shemu:
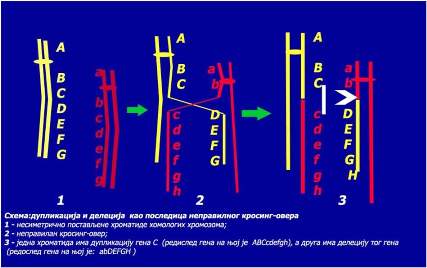
Duplikacije manje oštećuju fenotip nego delecije. Sa stanovišta evolucije
duplikacije su značajne jer dovode do usložnjavanja genetičkog materijala.
Delecije autosomnih hromozoma su uvek štetne, a ako su u homozigotnom stanju onda su letalne. Prvo opisano oboljenje uzrokovano delecijom je sindrom mačjeg plača. Ovaj sindrom nastaje kao posljedica delecije kratkog kraka hromozoma 5 (5r-). Naziv je dobio prema specifičnom oglašavanju beba, a praćen je umnom zaostalošću. Ako dođe do prekida na krajevima hromozoma tako da se izgube telomere, koje hromozomu daju stabilnost, onda prekinuti krajevi postaju lepljivi i međusobno se spajaju u jednu prstenastu strukturu. Takvi hromozomi se nazivaju ring-hromozomi. Poremećaj rasta je najizrazitija fenotipska osobina ove aberacije.
Translokacije
Translokacije predstavljaju razmjenu dijelova između homologih i nehomologih
hromozoma. Razlikuju se:
- Recipročne
- Nerecipročne
- Robertsonove translokacije
Pri recipročnoj translokaciji se razmjenjuju delovi između
bilo koja dva hromozoma.
Nerecipročna translokacija je prenos genetičkog materijala iz jednog u
drugi hromozom.
Robertsonove translokacije su razmjene dijelova između akrocentričnih
hromozoma, pri čemu ti hromozomi mogu biti isti (npr. 21/21) ili različiti
(npr. 14/21). Pri ovoj translokaciji dolazi do istovremenog prekida na
jednom hromozomu iznad, a na drugom ispod centromere. Zatim dolazi do
međusobnog spajanja prekinutih delova i to tako što se spoje dugi kraci
(q) oba hromozoma, a kratki kraci (r) se izgube u prvoj narednoj diobi.
Spajanjem dugih krakova oba hromozoma nastaje jedan metacentričan (ako
su učestvovali isti hromozomi) ili submetacentričan hromozom (ako su se
spojili q kraci različitih hromozoma).
Osobe nosioci translokacija su najčešće fenotipski normalne jer se gube
samo kratki kraci akrocentričnih hromozoma. (Kratki kraci sadrže gene
čiji nedostatak ne utiče na fenotip.) Za njih se kaže da su nosioci uravnotežene
translokacije. Potomstvo ovakvih osoba je ugroženo i može imati poremećen
broj hromozoma što izaziva određene sindrome (npr. Daunov sindrom). Takvi
potomci imaju neuravnoteženu translokaciju.
Potomstvo osoba koje su nosioci uravnotežene translokacije je u različitom
stepenu ugroženo u zavisnosti od toga da li se translokacija desila izmađu
istih ili različitih hromozoma. Kada je npr. u pitanju translokacija 14/21
onda takvi roditelji stvaraju i normalne gamete (imaju hromozome 14 i
21) i gamete koji nose translokaciju (imaju spojene hromozome 14/21) pa
mogu imati i normalnu i oboljelu decu. U slučaju translokacije 21/21 (između
istih hromozoma) osoba ne stvara normalne gamete već samo ili gamete koji
imaju oba hromozoma 21 (zakačeni su jedan za drugi) ili su bez hromozoma
2. Zbog toga oni ne mogu imati zdravu djecu.
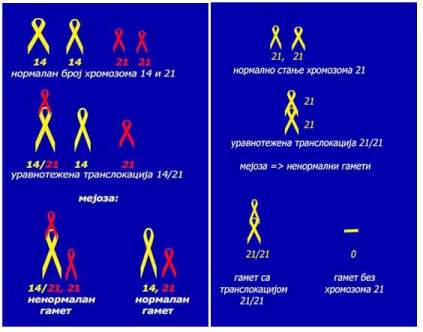
Uravnotežena translokacija 14/21 Uravnotežena translokacija 21/21
Inverzije
Inverzije nastaju kada se na jednom hromozomu dese dva prekida pa se
dio između prekida rotira za 180°, a zatim dolazi do prespajanja rotiranog
dijela za ostatak hromozoma. Time se mijenja redoslijed gena na hromozomu.
Inverzije mogu biti :
- Pericentrične
- Paracentrične
Ako je rotirani dio hromozoma obuhvatio centromeru onda je to pericentrična, a ako se rotacija desila samo unutar jednog kraka (nije zahvatila centromeru) onda je u pitanju paracentrična inverzija. Pericentričnim inverzijama dolazi do promjene oblika hromozoma. Nosioci inverzija su uglavnom fenotipski normalni. Štetnost inverzija izražava se u potomstvu. Zbog krosing-overa nastaju aberantni hromozomi koji se prenose na potomstvo.
Numeričke aberacije
Prema broju hromozoma nazivaju se numeričke aberacije. Razlikuju se dva
tipa ovih aberacija:
- Poliploidija
- Aneuploidija.
Poliploidija je numerička aberacija pri kojoj dolazi do uvećanja broja
ukupnih garnitura hromozoma. Ako se u tjelesnoj ćeliji nalazi više od
dvije, a u polnoj više od jedne garniture hromozoma onda su takve ćelije
poliploidne.
Tako nastaju ćelije sa triploidnim (3n), tetraploidnim (4n), pentaploidnim
(5n) itd. brojem hromozoma. Kod poliploidnih ćelija dolazi do uvećanja
osnovnog – monoploidnog broja hromozoma. Vrste koje nisu poliploidne u
tjelesnim ćelijama imaju dve garniture hromozoma (2n – diploidne su),
a u polnim ćelijama jednu garnituru (n – haploidan broj), tako da je kod
njih monoploidan broj jednak haploidnom broju hromozoma. Kod poliploidnih
vrsta se monoploidan i haploidan broj razlikuju. Uzroci poliploidija mogu
biti sledeći :
1. oplođenje jajne ćelije većim brojem spermatozoida, što predstavlja
najčešći uzrok pojave poliploidije kod čoveka;
2. oplođenje nenormalne jajne ćelije kod koje nije došlo do redukcije
broja hromozoma; ako se npr. oplodi diploidna jajna ćelija nastaće triploidan
zigot (2n + n= 3n);
3. spajanje diploidnih zigota, čime nastaje tetraploidan zigot (4n).
Poliploidija je vrlo česta i normalna pojava kod biljaka. U toku evolucije
cvijetnica smatra se da je oko 1/3 vrsta nastalo poliploidijom. Mnoge
kulturne vrste je čovek stvorio poliploidijom, jer su poliploidne biljke
krupnije, sa većim sadržajem hranljivih materija i dr. (npr. jagoda, banana,
šljiva, kruška, pšenica i dr.). Tako npr., već pomenuta, divlja hrizantema
ima sitan i neugledan cvet, dok je dekaploidna vrsta lepog, krupnog cveta.
Kod životinja je poliploidija rijetka kao normalna pojava; sreće se kod
vinske mušice, daždevnjaka, nižih račića i dr.
Za čovjeka je poliploidija štetna i uglavnom letalna i javlja se najčešće
kao:
- triploidija (3n) i
- tetraploidija (4n).
Jako se retko rađaju djeca sa ovim aberacijama jer se poliploidni zigoti
eliminišu još u intrauterinom razviću.
Aneuploidija
Aneuploidija predstavlja uvećanje ili smanjenje osnovne garniture za
jedan ili veći broj hromozoma. Uzroci nastanka ove aberacije su najčešće
:
1. nerazdvajanje parova hromozoma u mejozi I;
2. nerazdvajanje sestrinskih hromatida u mejozi II;
3. nerazdvajanje sestrinskih hromatida u mitozi.
Svaki hromozom se u normalnoj telesnoj ćeliji nalazi u paru pa se naziva
disomik (lat. di = dva; soma = telo).
Ako tjelesna ćelija umesto para hromozoma sadrži samo jedan onda je to
monosomik. Takva monosomična ćelija u ukupnom broju hromozoma ima jedan
hromozom manje od normalnog broja, pa je opšta formula monosomije 2n –1
(monosomična tjelesna ćelija čoveka ima 46 – 1 = 45 hromozoma).
Ćelija sa jednim hromozomom više od normalnog broja je trisomik (umesto
para određenog hromozoma ona ima tri) - opšta formula je 2n+1.
Može se desiti da ćelija ima dva hromozoma manje, odnosno da joj jedan
par hromozoma nedostaje pa je onda ta ćelija nultisomik(2n-2).
Ako, pak, ćelija ima dva hromozoma više od normalnog tj. umesto jednog
ima dva para određenog hromozoma, onda je ona tetrasomik(2n+2).
Pored ovih pobrojanih aberantnih stanja mogu se javiti i polisomije.
Nedostatak hromozoma je za ćeliju štetniji od viška hromozoma pri čemu
su monozomije autozomnih hromozoma letalne. Mada je letalnost aneuploidija
veoma velika neke od njih su prisutne u živorođenoj populaciji ljudi.
Takvi poremećaji najčešće imaju svojstva sindroma . Najčešća trizomija
koja se javlja u ljudskoj populaciji je trizomija 21, koja se fenotipski
izražava kao – Daunov sindrom
. Trizomija 13, se fenotipski izražava kao Patauov sindrom, je posle Daunovog
sindroma po učestalosti javljanja u ljudskoj populaciji (učestalost 1:5000).
Kariotip je 47,XX,+13 ili 47,XY,+13. Trizomija 18, fenotipski daje Edvardsov
sindrom, i ima kariotip 47,XX,+18 ili 47,XY,+18.
Aneuploidije polnih hromozoma manje oštećuju fenotip nego aneuploidije
autosomnih hromozoma. Osobe sa aneuploidijama autosomnih hromozoma pored
umne zaostalosti nose i niz drugih teških anomalija zbog kojih ne žive
dugo, dok je za osobe sa poremećenim brojem polnih hromozoma glavno obilježje
njihova sterilnost (neplodnost). Razlozi za to su što se na polnim hromozomima
nalazi manji broj gena nego na autozomnim i što svaki prekobrojni X hromozom
postaje neaktivan (to je u stvari tzv.Barovo tijelo).
Najčešće aneuploidije polnih hromozoma su:
- trisomija X hromozoma žene (kariotip je 47,XXX), nema naziv sindroma
jer su takve žene fenotipski normalne i klinički zdrave; neke imaju emotivne
teškoće u društvenom prilagođavanju;
- monosomija X hromozoma žene, koja fenotipski daje Tarnerov sindrom (45,XO)
čije je glavno obilježje sterilnost;
- disomija X hromozoma muškarca – Klinefelterov sindrom (47,XXY); radi
se o sterilnim muškarcima .
Kariotip je broj hromozoma u tjelesnim ćelijama koji je stalan i karakterističan za svaku biološku vrstu. Kariogram je grafički prikaz kariotipa pri čemu su hromozomi poredani po veličini.
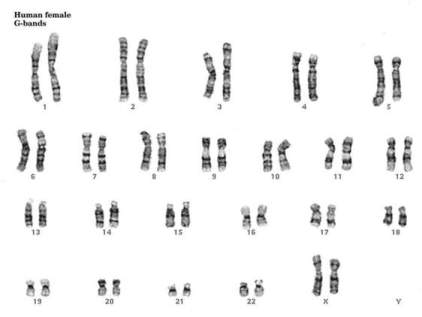
Kariogram žene
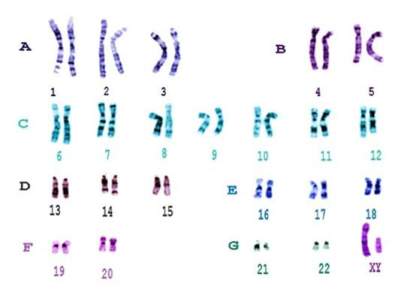
Kariogram muškarca
preuzmi
seminarski rad u wordu » » »
