OSTALI SEMINARSKI RADOVI
IZ BIOLOGIJE : |
|||||||||||||||||||||||||||||||||||||||||||
|
|||||||||||||||||||||||||||||||||||||||||||
POPRAVAK DNK
Tema o kojoj ćemo ovdje govoriti na prvi pogled izgleda vrlo složeno i nerazumljivo.Obilje literature i nepregledan materijal govore o njenoj važnosti. Budući da je DNK jedinstvena i trajna kopija genoma ona je vrlo bitna za zdravlje pojedinca i za uspješno održavanje reproduktivnosti. Međutim kao i sve druge molekule u našem organizmu i DNK u živoj stanici može biti izložena različitim hemijskim utjecajima. Na taj način dolazi do promjena u njenoj strukturi i dolazi do pojave mutacija. A znamo da bilo koja promjena u DNK sekvenci organizma predstavlja mutaciju. Sve stanice posjeduju enzime za DNK popravak koji pokušavaju smanjiti broj mutacija koje se događaju. Hemijske promjene DNK se događaju spontano ili pod utjecajem raznih hemikalija i zračenja. Da bi se popravila oštećena DNK, stanica je razvila različite mehanizme kao što su: izravni obrat oštećenja DNK, popravak izrezivanjem, popravak sklon pogreškama i rekombinacijski popravak. ( Cooper, Hausman, 2004 )
SISTEMI POPRAVKE DNK
 Na
hiljade genoma pati svaki dan zbog raznih oštećenja i zato je bitno da
stanica posjeduje učinkovit sistem za popravak. Bez popravka genom ne
bi bio u mogućnosti održavati svoje osnovne stanične funkcije više od
nekoliko sati. ( Brown, 2002 )
Na
hiljade genoma pati svaki dan zbog raznih oštećenja i zato je bitno da
stanica posjeduje učinkovit sistem za popravak. Bez popravka genom ne
bi bio u mogućnosti održavati svoje osnovne stanične funkcije više od
nekoliko sati. ( Brown, 2002 )
Većina stanica posjeduje 4 različita tipa popravke DNK
sistema (Lindahl, Wood, 1999 ) ( Slika 2.1.) :
• Izravni popravak oštećenja DNK, kao što samo ime govori,
djeluje direktno na oštećene nukleotide, vraćajući ih u izvornu strukturu.
• Popravak ekscizijom ( izrezivanjem nukleotida ) uključuje
odstranjenje jednog oštećenog segmenta nukleotida praćenog resintezom
ispravnog slijeda nukleotida od strane DNK polimeraze.
• Popravak krivo sparenih baza ispravlja pogreške napravljene
tokom replikacije tako što dolazi do isjecanja DNK između loma lanca i
krivo sparenih baza.
• Rekombinacijski popravak se zasniva na zamjeni oštećene
DNK rekombinacijom s neoštećenom molekulom.
Većina organizama, ako ne i svi, takođe posjeduju sisteme koji im omogućavaju
da repliciraju oštećena područja njihovog genoma bez prethodne popravke.
( Brown, 2002 )
2.1. IZRAVNI POPRAVAK OŠTEĆENJA DNK
Većina oštećenja DNK, nastala pod utjecajem hemijskih ili fizičkih mutagena,
može se popraviti rezanjem oštećenih nukleotida nakon čega slijedi resinteza
ispružene DNK kao što je prikazano na slici 2.1. Samo se nekoliko vrsta
oštećenih nukleotida može popraviti izravno ( Brown, 2002 ) :
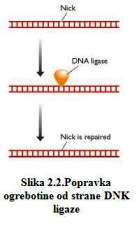 •
Ogrebotine može popraviti DNK ligaza samo ukoliko je fosfodiesterska veza
pukla i ukoliko nema oštećenja na 5'- fosfatnoj i 3' – hidroksilnoj grupi
nukleotida,na obje strane šupljine. To je često slučaj sa ogrebotinama
nastalim djelovanjem jonizirajućeg zračenja. ( Slika 2.2. )
•
Ogrebotine može popraviti DNK ligaza samo ukoliko je fosfodiesterska veza
pukla i ukoliko nema oštećenja na 5'- fosfatnoj i 3' – hidroksilnoj grupi
nukleotida,na obje strane šupljine. To je često slučaj sa ogrebotinama
nastalim djelovanjem jonizirajućeg zračenja. ( Slika 2.2. )
• Neki oblici alkilirajućih oštećenja se mogu direktno popraviti reverzibilnim
enzimima koji prenose alkilnu grupu od nukleotida do vlastitih polipeptidnih
lanaca.Enzimi sposobni za ovaj popravak se nalaze u mnogim organizmima
i tu npr. spada Ada enzim E. coli koji je uključen u proces prilagodbe,
tako da ga ova bakterija može aktivirati kao odgovor na oštećenje DNK.
Ada uklanja alkilne grupe zakačene na kisikove grupe na poziciji 4 i 6
timina i guanina, a takođe može popraviti fosfodiesterske veze koje su
postale metilirane. Međutim neki enzimi imaju ograničeno djelovanje, primjer
je ljudski MGMT (O6-metilguanin-DNA metiltransferaza ) koji , kako mu
samo ime govori, samo uklanja alkilne grupe sa pozicije 6 guanina.
• Ciklobutilni dimeri se saniraju putem svjetla odnosno procesom fotoreaktivacije.
Kod E. coli ovaj proces uključuje enzim DNK fotoliazu. Kada je stimulirana
svjetlom valne dužine između 300 i 500 nm, enzim se veže za ciklobutilne
dimere i vraća ih u prvobitne monomerne nukleotide. Fotoreaktivacija je
široko rasprostranjena međutim ona nije univerzalna vrsta popravke: poznata
je kod mnogih , ali ne kod svih bakterija, dosta kod eukariota uključujući
i neke kičmenjake, ali je nema kod ljudi i drugih sisara sa placentom.
2.2. POPRAVAK EKSCIZIJOM
 Izravne
popravke oštećenja DNK opisane gore su važne , ali one čine vrlo malu
komponentu među sistemima popravke DNK većine organizama.Ovo je zbog toga
što nacrt sekvenci ljudskog genoma sadrži samo jedan gen za kodiranje
proteina uključenih u direktnu popravku ( MGMT gen ), ali koji ima najmanje
40 gena za puteve ekscizijskog popravka ( Wood et. Al., 2001 ).
Izravne
popravke oštećenja DNK opisane gore su važne , ali one čine vrlo malu
komponentu među sistemima popravke DNK većine organizama.Ovo je zbog toga
što nacrt sekvenci ljudskog genoma sadrži samo jedan gen za kodiranje
proteina uključenih u direktnu popravku ( MGMT gen ), ali koji ima najmanje
40 gena za puteve ekscizijskog popravka ( Wood et. Al., 2001 ).
Puteve ekscizijskog popravka možemo podijeliti u dvije kategorije ( Brown,
2002 ) :
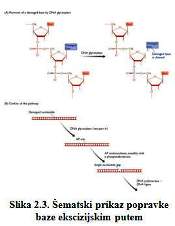
• Popravak izrezivanjem baza uključuje uklanjanje oštećene
baze, izrezivanjem kratkog komada polinukleotida oko AP mjesta, i ponovnu
resintezu uz pomoć DNK polimeraze. ( Slika 2.3. )
• Popravak izrezivanjem nukleotida je sličan prethodno
opisanom popravku , međutim njemu ne prethodi uklanjanje oštećene baze
i može djelovati na znatno više područja oštećene DNK.
Sada ćemo objasniti svaki od ova dva puta redom.
Popravak izrezivanjem baza sanira mnoge vrste oštećenih
nukleotida. On je najmanje složen za razliku od raznih sistema popravka
koji uključuju uklanjanje jednog ili više oštećenih nukleotida i resintezu
DNK kako bi obuhvatila nastalu prazninu. Koristi se za popravak mnogih
modificiranih nukleotida čije su baze pretrpjele manju štetu koja je rezultat
, naprimjer, izloženosti alkilirajućim agensima ili jonizirajućem zračenju.
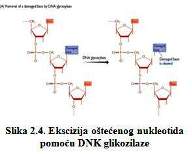
Proces se pokreće od strane DNK glikozilaze koja prione na β-N-glikozidnu
vezu između oštećene baze i šećerne komponente nukleotida ( Slika 2.4.).
Svaka DNK glikozilaza ima ograničenu specifičnost ( Brown, 2002 ).
 Mnogi organizmi se mogu nositi sa dezaminiranim bazama kao što su uracil
( dezaminirani citozin ) i hipoksantin ( dezaminirani adenin ), zatim
sa oksidacionim produktima kao što su 5 – hidroksicitozin i timin glikol,
i sa metiliranim bazama kao što su 3 – metiladenin, 7 – metilguanin i
2 – metilcitozin ( Slika 2.5. ) ( Seeberg et. al., 1995 )
Mnogi organizmi se mogu nositi sa dezaminiranim bazama kao što su uracil
( dezaminirani citozin ) i hipoksantin ( dezaminirani adenin ), zatim
sa oksidacionim produktima kao što su 5 – hidroksicitozin i timin glikol,
i sa metiliranim bazama kao što su 3 – metiladenin, 7 – metilguanin i
2 – metilcitozin ( Slika 2.5. ) ( Seeberg et. al., 1995 )
Ostale DNK glikozilaze uklanjaju normalne baze u sistemu popravke krivo
sparenih baza.
DNK glikozilaza uklanja oštećenu bazu preklapanjem struktura na mjestu
izvan heliksa, a zatim je odvaja od polinukleotida ( Kunkel, Wilson, 1996
; Roberts, Cheng, 1998 ). Tako se stvara AP mjesto koje se pretvara u
jedan nukleotidni otvor u drugom koraku popravke. Ovaj korak se može obavljati
na razne načine. Standardna metoda koristi AP – endonukleazu kao što su
endonukleaza III ili endonukleaza IV E. coli ili ljudski APE1 koji presjeca
fosfodiestersku vezu na 5' kraju AP mjesta.Neke AP – endonukleaze mogu
ukloniti šećer iz AP mjesta. Alternativni put, za pretvaranje AP mjesta
u otvor, koristi endonukleaznu aktivnost koju imaju neke DNK glikozilaze,
koje mogu napraviti rez na 3' kraju AP mjesta, vjerovatno u isto vrijeme
kada se oštećena baza ukloni.To je sve praćeno odvajanjem šećera od strane
fosfodiesteraze ( Brown, 2002 ).
Nukleotidni otvor popunjavaju DNK polimeraze koristeći se uparivanjem
baze sa neoštećenom bazom na drugoj strani molekule DNK kako bi bilo sigurno
da je tačan nukleotid umetnut. Kod E. coli otvor je popunjen DNK polimerazom
I , a kod sisara DNK polimerazom β ( Sobol et. al., 1996 ). Kvasac koristi
DNK replicirajući enzim, DNK polimerazu σ, za ovu svrhu ( Seeberg et.
al., 1995. ) Nakon popunjavanja praznine, konačno fosfodiesterska veza
je na svom mjestu uz pomoć DNK ligaze ( Brown, 2002 ).
Popravak izrezivanjem nukleotida se koristi za saniranje mnogo većih vrsta
oštećenja zato što ima puno više specifičan sistem popravka i u mogućnosti
je da se nosi sa ekstremnim oblicima oštećenja kao što je npr. ispravljanje
ciklobutilnih dimera sistemom tamnog popravka pod uslovom da ti organizmi
nemaju fotoreaktivni sistem ( kao ljudi ) . Kod popravka izrezivanjem
nukleotida , segment jednog lanca DNK sadrži oštećene nukleotide i on
se izreže i zamijeni s novom DNK. Proces je zbog ovoga sličan popravku
izrezivanjem baze jedino je razlika što mu ne prethodi selektivno uklanjanje
baze i dugi polinukleotid je izrezan. Najbolje istražen primjer popravke
izrezivanjem nukleotida je kratki proces zakrpe E. coli , tako se zove
zato što je regija polinukleotida, izrezana i naknadno zakrpljena, relativno
kratka, obično sadrži 12 nuklotida po dužini. Popravak sistemom kratke
zakrpe je pokrenut uz pomoć multienzimskog kompleksa koji se zove UvrABC
endonukleaza. U prvoj fazi procesa trimer se sastoji od dva UvrA proteina,
a jedna kopija UvrB se kači na oštećeno mjesto DNK. Široka specifičnost
procesa pokazuje da pojedine vrste oštećenja nisu izravno otkrivene i
da kompleks mora puno više istražiti osobine oštećenja DNK kao što je
narušavanje dvostruke spirale ( heliksa ).UvrA je dio kompleksa koji je
možda najviše uključen na mjestu oštećenja zato što disocira u trenutku
pronalska položaja oštećenja i ne igra nikakvu ulogu u daljenjem postupku
popravke. Odlazak UvrA omogućuje UvrC- u povezivanje , stvara se UvrBC
dimer koji sječe polinukleotid na obje strane oštećenja. Prvi rez pravi
UvrB na petoj fosfodiesterskoj vezi nizvodno od oštećenog nukleotida,
a drugi rez pravi UvrC na osmoj fosfodiesterskoj vezi uzvodno. Kao rezultat
dobivamo izrezanih 12 nukleotida, iako postoje neke varijabilnosti, pogotovo
na mjestu UvrB reza. Izrezani segment se zatim uklanja obično kao oligonukleotid
od strane DNK helikaze II koja vjerovatno odvoji segment tako što razbije
bazne parove. UvrC se takođe odvaja u ovoj fazi, ali UvrB ostaje na mjestu
i premošćuje otvor nastao rezanjem. Uloga UvrB – a je da spriječi uparivanje
baza samih sa sobom jednog dijela jednloančane DNK, a njegova alternatina
uloga je da spriječi ovaj lanac od oštećenja ili da eventualno usmjeri
DNK polimerazu na mjesto koje treba popraviti.Otvor popunjava DNK polimeraza
I i posljednju fosfodiestersku vezu sintetizira DNK ligaza ( Brown, 2002
).
E. coli ima sistem duge zakrpe, popravke izrezivanjem nuklotida, koji
uključuje Uvr proteine, ali je razlika u tome da dio izrezane DNK može
biti veći od 2 kb po dužini. Ovaj sistem je manje proučavan i proces se
dobro ne razumije, ali pretpostavlja se ima ulogu kod većih oštećenja,
u regijama skupine nukleotida a ne kod pojedinačnih ( Brown, 2002 ).
Kod eukariotskih nukleotida ekscizijski proces se takođe zove sistem duge
zakrpe, ali kao rezultat imamo zamjenu od 24 – 29 nukleotida DNK. Nema
kod eukariota sistema kratke zakrpe, a ime se koristi za razlikovanje
ovog procesa od popravke izrezivanjem baza. Sistem je složeniji nego kod
E. coli i čini se da ti relevantni enzimi nisu homolozi Uvr proteina.
Kod ljudi najmanje 16 proteina je uključeno, sa jednim rezom nizvodno
napravljenim na istom mjestu kao kod E. coli – peta fosfodiesterska veza
– ali sa više udaljenim uzvodnim rezom, što rezultira dužim rezanjem.Oba
reza su napravljena od strane endonukleaze ( Brown, 2002 ).
Ova aktivnost je osigurana djelimično TFIIH, jednim od sastavnih dijelova
RNK polimeraze II inicijacijskog kompleksa. Isprva se pretpostavljalo
da TFIIH ima dvostruku ulogu u ćeliji, funkcionirajući odvojeno u transkripciji
i sistemu popravke, ali sada se smatra da postoji izravna veza između
ova dva procesa ( Lehmann, 1995; Svejstrup et. al., 1996 ). Ovaj stav
je podržan otkrićem popravka povezanog sa transkripcijom, koji je specifičan
za popravak oštećenja unutar gena koji se prepisuju. Prvi tip ovog popravka
je modificirana verzija ekscizije nukleotida, ali sada je poznato da je
i bazna ekscizija povezana sa transkripcijom ( Cooper et. al., 1997 ).
Ova otkrića ne znače da netranskribirajući regioni genoma nisu sanirani.Ekscizijski
proces popravke štiti cijeli genom od oštećenja, ali je sasvim logično
da posebni mehanizmi trebaju postojati radi usmjeravanja procesa i gena
koji se prepišu. Gen sadrži biološku informaciju genoma i održavanje njegovog
integriteta bi trebao biti najveći prioritet sistemima popravke ( Brown,
2002 ) .
2.3. POPRAVAK KRIVO SPARENIH BAZA
Popravak krivo sparenih baza ispravlja pogreške napravljene tokom replikacije.Svaki
od sistema popravka koje smo gore naveli ( izravni popravak, ekscizija
baze i nukleotida ), prepoznaju i djeluju na oštećenje DNK uzrokovano
mutagenima. To znači da oni traže abnormalne hemijske strukture poput
modificiranih nukleotida i ciklobutilnih dimera.Međutim oni ne mogu popraviti
krivo sparene baze koje su rezultat grešaka u replikaciji. Popravak krivo
sparenih baza ispravlja replikacijske greške, ali on ne detektuje krivo
sparene nukleotide nego detektuje nedostatak baznih parova između roditelja
i kćerke ćelije. Kada se krivo sparene baze pronađu, ovaj sistem popravke
reže dio polinukleotida kćerke ćelije i ispunjava praznine, na sličan
način kao kod bazne i nukleotidne ekscizije.Popravak se mora desiti u
polinukleotidu kćerke ćelije, jer je u ovom novosintetiziranom lancu došlo
do pogreške; polinukleotid roditelja ima pravilan slijed ( sekvencu ).
Kako sistem popravka zna koji je lanac čiji? Kod E. coli kćerkin lanac
je slabo metiliran te se može razlikovati od roditeljskog polinukleotidnog
lanca, koji ima puni set metilnih grupa. Kod E. coli DNK je metilirana
zbog djelovanja DNK adenin metilaze ( Dam ), koja konvertuje adenin u
6 – metiladenin u slijedu 5' – GATC – 3' i DNK citozin metilaze ( Dcm
) , koja konvertuje citozin u 5 – metilcitozin u slijedu 5' – CCAGG –
3' i 5' – CCTGG – 3'. Ove metilacije nisu mutageni, modificirani nukleotidi
imaju osobine istog baznog uparivanja kao nepromijenjene verzije. Postoji
kašenjenje kod DNK replikacije i metilacije kćerkinog lanca i tokom tog
kašenjenja postoji prilika da sistem popravka skenira krivo sparene baze
i da učini potrebne ispravke u slabo metiliranom kćerkinom lancu ( Slika
2.8. ) ( Brown, 2002 ).
E. coli ima 3 sistema za popravak krivo sparenih baza: „ duga zakrpa“,
„ kratka zakrpa“ i „ vrlo kratka zakrpa“. Imena pokazuju relativne dužine
izrezanih i resinteziranih segmenata. Dugi sistem zakrpe zamjenjuje do
kb ili više DNK i zahtijeva MutH, MutL i MutS proteine kao i DNK helikazu
II koju smo susreli tokom nukleotidne ekscizije. MutS prepoznaje krivo
sparene baze, a MutH razlikuje dva lanca vezujući se za nemetilirane 5'
– GATC – 3' sekvence ( Brown, 2002 ).
Uloga MutL je nejasna, ali bi on mogao koordinirati aktivnosti druga
dva proteina tako da se MutH veže za 5' – GATC – 3' sekvencu samo u blizini
mjesta krivo sparenih baza koje prepoznaje MutS. Nakon vezanja, MutH reže
fosfodiestersku vezu uzvodno od G u metilacijskoj sekvenci i DNK helikaza
II odvaja jedan lanac. Ne javlja se enzim koji reže lanac nizvodno od
mjesta krivo sparenih baza; umjesto toga usamljena jednolančana regija
biva degradirana od strane egzonukleaze koja prati helikazu i dalje nastavlja
da se kreće izvan krivo sparenih baza. Otvor popunjava DNK polimeraza
I i DNK ligaza. Slično se događa kod kratkog i vrlo kratkog popravka krivo
sparenih baza, jedino je razlika u specifičnosti proteina koji prepoznaju
neusklađenost. Kratki sistem popravke, koji je rezultat rezanja segmenta
manje od 10 nukleotida po dužini, počinje kada MutY prepoznaje A – G ili
A – C neusklađenost, a vrlo kratki sistem popravka ispravlja G – T neusklađenosti
koje su prepoznate od strane Vsr endonukleaze ( Brown, 2002 ).
Eukarioti imaju homologe Mut proteina E. coli i njihov proces popravka
krivo sparenih baza vjerovatno radi na sličan način ( Kolodner, 2000 ).
Jedina razlika je u tome da metilacija neće biti metod koji se koristi
za razlikovanje između roditeljskog i kćerkinog polinukleotidnog lanca.
Metilacija je upletena u popravak krivo sparenih baza u stanicama sisara,
ali i kod DNK nekih eukariota, uključujući voćne mušice i kvasac, koja
nije puno metilirana ( Brown, 2002 ).
2.4. Rekombinacijski popravak
Bez rekombinacije , genomi bi bili relativno statične strukture, podložne
vrlo malim promjenama. Postupno nakupljanje mutacija tokom dužeg vremenskog
razdoblja bi rezultiralo malim promjenama u slijedu nukleotida genoma,
ali mnogo opsežnijem restruktiriranjem, što je uloga rekombinacije, ali
to neće nastupiti i evolucijski potencijal genoma bi bio strogo ograničen
( Brown, 2002 ).
Rekombinacija je prvi put prepoznata kao proces odgovoran za crossing
– over i izmjenu DNK segmenata između homolognih hromosoma tokom mejoze
u eukariotskim stanicama, a potom je upletena u integraciju prenosa DNK
u bakterijski genom nakon konjugacije, transdukcije ili transformacije
( Brown, 2002 ).
Biološkom važnošću ovih procesa došlo je do prvih pokušaja da se opišu
molekularni događaji koji su uključeni u rekombinaciju i tako je došlo
do otkrića Hollidayevog modela ( Holliday, 1964 ), s kojim ćemo započeti
priču o rekombinaciji.
Homologna rekombinacija
Hollidayev model se odnosi na vrstu rekombinacije koja se zove opća ili homologna rekombinacija. Ovo je najvažniji tip rekombinacije u prirodi i odgovorna je mejotski crossing – over i za integraciju prenešene DNK u bakterijskom genomu ( Brown , 2002 ).
Hollidayev model homologne rekombinacije
Hollidayev model opisuje rekombinaciju između dvije homologne molekule, sa istim ili gotovo identičnim sekvencama. Najvažniji značaj modela je formacija heterodupleksa koji je rezultat razmjene polinukleotidnih segmenata između dvije homologne molekule.
Heterodupleks je na početku stabiliziran baznim uparivanjem između svakog
prenesenog lanca i netaknutim polinukeotidom receptorne molekule . Ovo
bazno uparivanje je moguće zbog sličnosti sekvenci između ove dvije molekule.
Nakon toga otvore začepi DNK ligaza, dajući Hollidayevu strukturu. Ova
struktura je vrlo dinamična.Do razrješavanja Hollidayeve strukture u individulane
molekule dolazi cijepanjem ukriženih lanaca. Ovo je ključ cijelog procesa,
jer rez može biti napravljen na bilo kojem od dva smjera. Ova dva reza
imaju različite rezultate. Ako je rez napravljen od lijeva na desno prelazi
hi oblik, sve što se dešava jeste da se kratki segment polinukleotida
prenosi između dvije molekule. S druge strane, rez napravljen u pravcu
gore – dole rezultuje uzajamnom razmjenom, dvolančana DNK se prenosi između
dvije molekule tako da se kraj jedne molekule promjeni za kraj druge molekule.To
je prenos DNK viđen u crossing – overu ( Brown, 2002 ).
Do sada smo ignorirali jedan aspekt Hollidayevog modela, a to je način
na koji su dvije dvolančane molekule na početku procesa djelovale da bi
nastao heterodupleks. U originalnoj shemi, dvije molekule su postavljene
jedan uz drugu i javlja je jednolančani urez na odgovarajućim pozicijama
u svakoj spirali. Tako se proizvode slobodni jednolančani krajevi koji
mogu biti promijenjeni, kao rezultat nastaje heterodupleks ( Brown, 2002
).
Značaj ovog modela je bio kritiziran, jer ne postoji mehanizam koji bi
tvrdio da su se urezi dogodili na tačno istom položaju svake molekule.
Meselson – Radding modifikacija ( Meselson, Radding, 1975 ) predlaže više
zadovoljavajuću shemu u kojoj se jednolančani urez javlja u samo jednom
od dvije spirale ( heliksa ), slobodni kraj, koji je proizveden napadom
na neprekinutu dvostruku zavojnicu na homolognom položaju , se razriješuje
i dolazi do formiranja D – petlje ( D – loop ) ( Brown, 2002 ).
Nakon cijepanja lanaca na spoju između jednolančane i bazne regije proizvodi
se heterodupleks.
Proteini uključeni u homolognu rekombinaciju kod E. coli
Hollidayev model i Meselson – Radding modifikacija odnose se na homologne
rekombinacije u svim organizmima, ali kao i u mnogim drugim područjima
molekularne biologije, početni napredak u razumijevanju kako se proces
provodi u ćeliji je napravljen kod E. coli. Specifični rekombinacijski
sistem, koji je proučavan, imao je kružni genom E. coli kao jednog partnera
i linearni hormosomski fragment kao drugog partnera.Ova situacija se javlja
tokom konjugacije, transdukcije ili transformacije bakterijskih stanica
( Brown, 2002 ).
Mutacijska istraživanja su identificirala broj gena E. coli , koji kada
se inaktiviraju dovode do grešaka u homolognoj rekombinaciji, što pokazuje
da su njihovi proteinski proizvodi uključeni u procese na neki način.
Opisana su 3 različita sistema rekombinacije, RecBCD, RecE, RecF putevi.
RecBCD je najvažniji kod bakterija ( Camerini – Otero, Hsieh, 1995 ).Kod
ovog puta , rekombinacija je pokrenuta od strane enzima RecBCD koji ima
nukleaznu u helikaznu aktivnost. Njegov precizan način djelovanja je neizvjestan:
u najjednostavnijem modelu enzim se veže na jednom kraju linearne molekule
i pokriva je dok ne dođe do prve kopije konsenzusa od 8 nukleotida sekvence
5' – GCTGGTGG – 3', koja se pojavljuje svakih 6 kb kod DNK E. coli ( Blattner
et. al., 1997 ). Nukleazna aktivnost enzima napravi jednolančani urez
na poziciji oko 56 nukeotida do 3' kraja „Hi“ strane ( Brown, 2002 ).
Neki kažu kažu da RecBCD enzim pravi ogrebotine dok napreduje uzduž linearne
DNK, ali ova aktivnost biva inhibirana kada dospije do kraja „ Hi „ strane.
Ove progresivno napravljene ogrebotine su ekvivalent jednoj ogrebotini
u prvom modelu ( Eggleston, West, 1996 ).
Bez obzira na precizan mehanizam, enzim RecBCD proizvodi slobodni jednolančani
kraj, koji prema Meselson – Radding modifikaciji , napada netaknutog partnera,
u ovom slučaju kružni genom E. coli . Posrednik ove faze je RecA protein,
koji formira proteinom – obloženi DNK filament koji može upasti u netaknutu
dvostruku spiralu i napraviti D – petlju ( D – loop ) ( Brown, 2002 ).
Posrednik u formiranju D – petlje je vjerovatno tripleks struktura, trolančana
DNK spirala u koju upada polinukleotid koji leži sa glavnim utorom netaknute
spirale i tvori vodikove veze sa baznim parovima koje susreće ( Camerini
– Otero, Hsieh, 1995 ).
ZAKLJUČAK
DNK je izložena različitim hemijskim reakcijama i promjena njene strukture bi dovela do različitih posljedica kao što su mutacije. Mutacije mogu nastati kao posljedica ugradnje pogrešne baze za vrijeme umnožavanja DNK. Izloženost DNK različitim hemikalijama ili zraženju takođe može blokirati replikaciju ili transkripciju. Da bi održale svoj genom stanice su razvile mehanizme za popravak oštećenja DNK, a to su: izravni popravak oštećenja DNK, popravak ekscizijom, popravak krivo sparenih baza i rekombinacijski popravak. Izravni popravak oštećenja DNK djeluje direktno na oštećene nukleotide, vraćajući ih u izvornu strukturu. U izravnom popravku oštećenja DNK ogrebotine nastale djelovanjem jonizirajućeg zračenja može popraviti DNK ligaza, alkilirajuća oštećenja se direktno popravljaju reverzibilnim enzimima kao što je npr. Ada enzim E.coli . Međutim neki enzimi imaju ograničeno djelovanje kao što je ljudski MGMT (O6-metilguanin-DNA metiltransferaza ) , koji uklanja samo alkilne grupe sa pozicije 6 guanina. Ciklobutilni dimeri se saniraju procesom fotoreaktivacije. Popravak ekscizijom ( izrezivanjem ) se može podijeliti na popravak izrezivanjem baza ( uklanjaju se oštećene baze ) i popravak izrezivanjem nukleotida ( njemu ne prethodi uklanjanje oštećene baze) . Kod izrezivanja baza DNK glikozilaza uklanja oštećenu bazu i taj postupak koristi se za popravak mnogih modificiranih nukleotida čije su baze pretrpjele manju štetu koja je rezultat izloženosti alkilirajućim agensima ili jonizirajućem zračenju.Popravak izrezivanjem nukleotida se koristi za saniranje mnogo većih vrsta oštećenja. Popravak krivo sparenih baza ispravlja krivo sparene baze koje su rezultat pogreške napravljene tokom replikacije. On ne detektuje krivo sparene nukleotide nego detektuje nedostatak baznih parova između roditelja i kćerke ćelije. Rekombinacijski popravak podrazumijeva zamjenu oštećene DNK rekombinacijom s neoštećenom molekulom. Ovajs sistem popravka se koristi za saniranje oštećenja za vrijeme replikacije DNK. Postoji opći ili generalni tip rekombinacije (odgovoran je za mejotski crossing – over i za integraciju prenešene DNK u bakterijskom genomu) i Hollidayev model homologne rekombinacije (opisuje rekombinaciju između dvije homologne molekule, sa istim ili gotovo identičnim sekvencama). Složeniji organizmi sa složenijim genomima imaju složenije mehanizme za popravak.Veliki broj proteina katalizira hemijske reakcije i igraju značajnu ulogu u mehanizmima popravka. Popravak DNK sklon pogreškama je uključen u procese koji smanjuju: ubijanje stanice, mutacije, replikaciju grešaka, ustrajnost DNK oštećenja i genomske nestabilnosti.Abnormalnosti u ovim procesima su vezani za pojavu raka i starenje.
SAŽETAK ( kratak opis rada )
U ovom seminarskom radu smo obradili načine na koje se popravlja oštećena DNK. DNK je sklona različitim napadima iz okoliša kao što je npr. rak kože, kojeg uzrokuju UV – zrake. Budući da je DNK spremište genetske informacije u svakoj živoj stanici, njen integritet i stabilnost su neophodni za život.Ako se DNK ne bi popravila to bi dovelo do mutacija i eventualnih bolesti. Procesi DNK popravka postoje i u prokariotskim i u eukariotskim organizmima i mnogi proteini su uključeni u taj popravak. Stanice su razvile čitav niz mehanizama da bi detektovale i popravile različite vrste oštećenja koje mogu nastati u DNK, bez obzira da li su te štete uzrokovane iz različitog okoliša ili su posljedica pogreški u replikaciji. Postoje 4 mehanizma kojim se vrši popravka DNK koje smo obradili u seminarskom radu, a to su: izravan popravak oštećenja DNK, popravak ekscizijom, popravak krivo sparenih baza i rekombinacijski popravak.
LITERATURA
1. Cooper, M.G., Hausman, E.R. ( 2004 ). Stanica – molekularni
pristup ( treće izdanje ). Zagreb: Medicinska naklada
2. Brown, T.A. ( 2002 ). Genomes. 2nd edition. Oxford: Willey - Liss
3. Lindahl, T., Wood, R.D. ( 1999 ). Quality control by DNA repair. Science.
PubMed: ;286:1897–1905.
4. Wood, R.D., Mitchell, M., Sgouros, J., Lindahl, T. ( 2001 ). Human
DNA repair genes. Science. PubMed: 291:1284–1289.
5. Seeberg, E. Eide, L., Bjoras, M. ( 1995 ). The base excision repair
pathway. Trends Biochem. Sci. PubMed: 20:391–397.
6. Kunkel, T.A., Wilson, S.H. ( 1996 ) . Push and pull of base flipping.
Nature. PubMed: 384:25–26.
7. Roberts, R.J., Cheng, X. ( 1998 ). Base flipping. Ann. Rev. Biochem.
PubMed: 67:181–198.
8. Sobol, R.W., Horton, J.K., Kuhn, R. et. al. ( 1996 ). Requirement of
mammalian DNA polymerase- β in base excision repair.Nature. PubMed: 379:183–186.
9. Lehmann,A.R.(1995). Nucleotide excision repair and the link with transcription.
Trends. Biochem.Sci. PubMed: 20:402–405.
10. Svejstrup, J.Q., Vichi, P., Egly, J-M. ( 1996 ). The multiple roles
of transcription factor / repair factor TFIIH. Trends. Biochem. Sci. PubMed:
21:346–350
11. Cooper, P.K., Nouspikel, T., Clarkson, S.G., Leadon, S.A. ( 1997 ).
Defective transcription – coupled repair of oxidative base damage in Cockayne
syndrome patients from XP group G. Science. PubMed: 275:990–993.
12. Kolodner, R.D.E. ( 2000 ). Guarding against mutation. Nature. PubMed:
407:687–689.
13. Holliday, R. ( 1964 ). A mechanism for gene conversion in fungi. Genet.
Res. PubMed: 5:282–304.
14. Meselson, M. , Radding, C.M. ( 1975 ). A general model for genetic
recombination. Proc. Natl Acad. Sci. USA. PubMed: 72:358–361
15. Camerini – Otero, R.D., Hsieh, P. ( 1995 ). Homologous recombination
proteins in prokaryotes and eukaryotes. Ann. Rev. Genet. PubMed: 29:509–552.
16. Blattner, F.R., Plunkett, G. Bloch, C.A. et. al. ( 1997 ). The complete
genome sequence of E.coli K-12. Science. PubMed: 277:1453–1462.
preuzmi
seminarski rad u wordu » » »

Besplatni Seminarski
Radovi